Såvitt man kan förstå är människans stamtavla mest utförligt beskriven i klassificeringen Natura Biota (Archaea Proteoarchaeota Eukaryomorpha) + (Bacteria Proteobacteria α-Proteobacteria) → Eukaryota Orthokaryota Neokaryota Scotokaryota Podiata Unikonta Obazoa Opisthokonta Holozoa Filozoa Choanozoa Animalia Eumetazoa Parahoxozoa Bilateria Nephrozoa Deuterostomia Chordata Olfactores Craniata Vertebrata Gnathostomata Eugnathostomata Euteleostomi Sarcopterygii Rhipidistia Tetrapodomorpha Eotetrapodiformes Elpistostegalia Stegocephalia Tetrapoda Reptiliomorpha Amniota Synapsida Eupelycosauria Metopophora Haptodontiformes Sphenacomorpha Sphenacodontia Pantherapsida Sphenacodontoidea Therapsida Eutherapsida Neotherapsida Theriodontia Eutheriodontia Cynodontia Epicynodontia Eucynodontia Probainognathia Chiniquodontoidea Prozostrodontia Mammaliamorpha Mammaliaformes Mammalia Theriiformes Holotheria Trechnotheria Cladotheria Zatheria Tribosphenida Theria Eutheria Placentalia Boreoeutheria Euarchontoglires Euarchonta Primatomorpha Primates Haplorhini Simiiformes Catarrhini Hominoidea Hominidae Homininae Hominini Hominina Homo sapiens sapiens, i enlighet med den systematik som fanns upp av Carl von Linné.
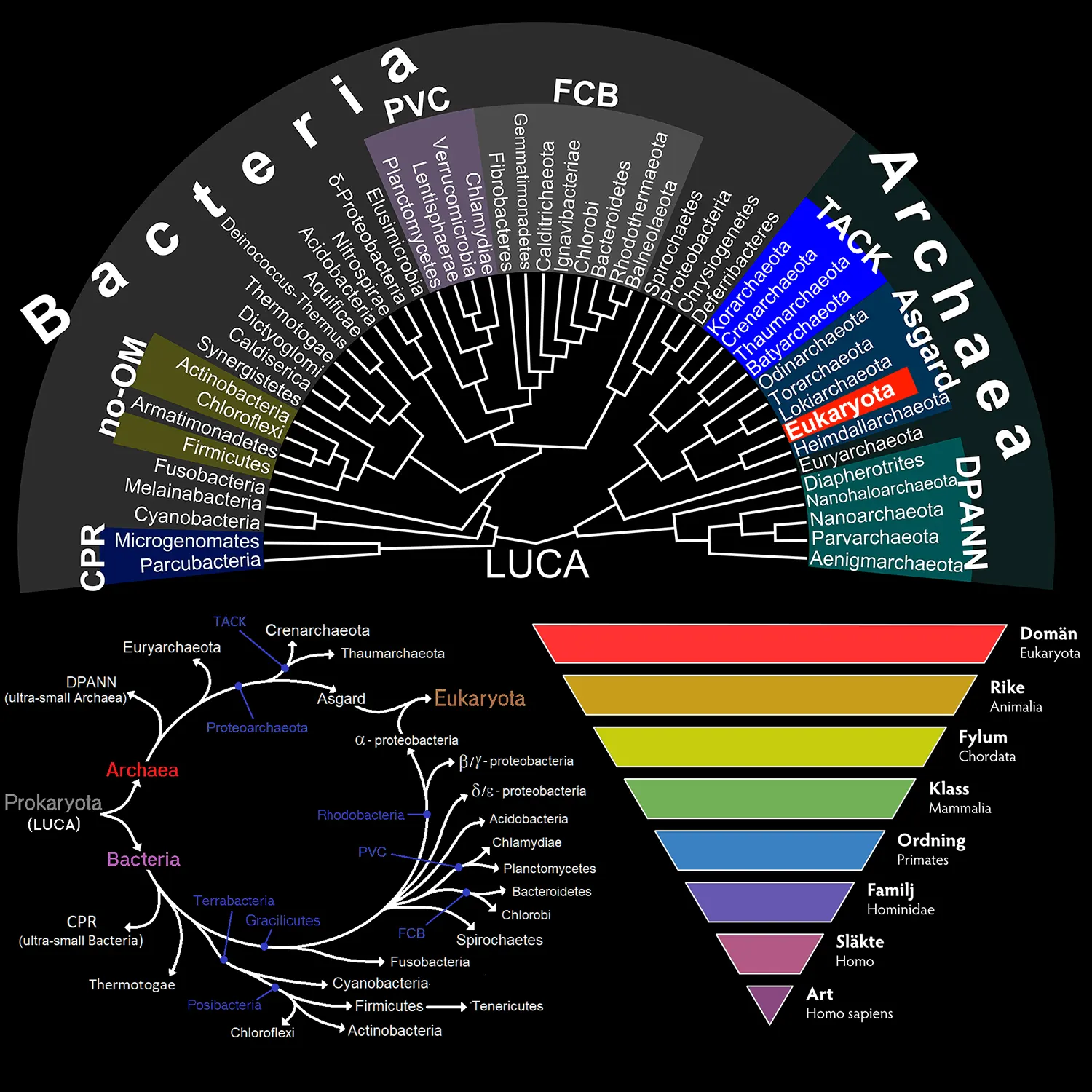
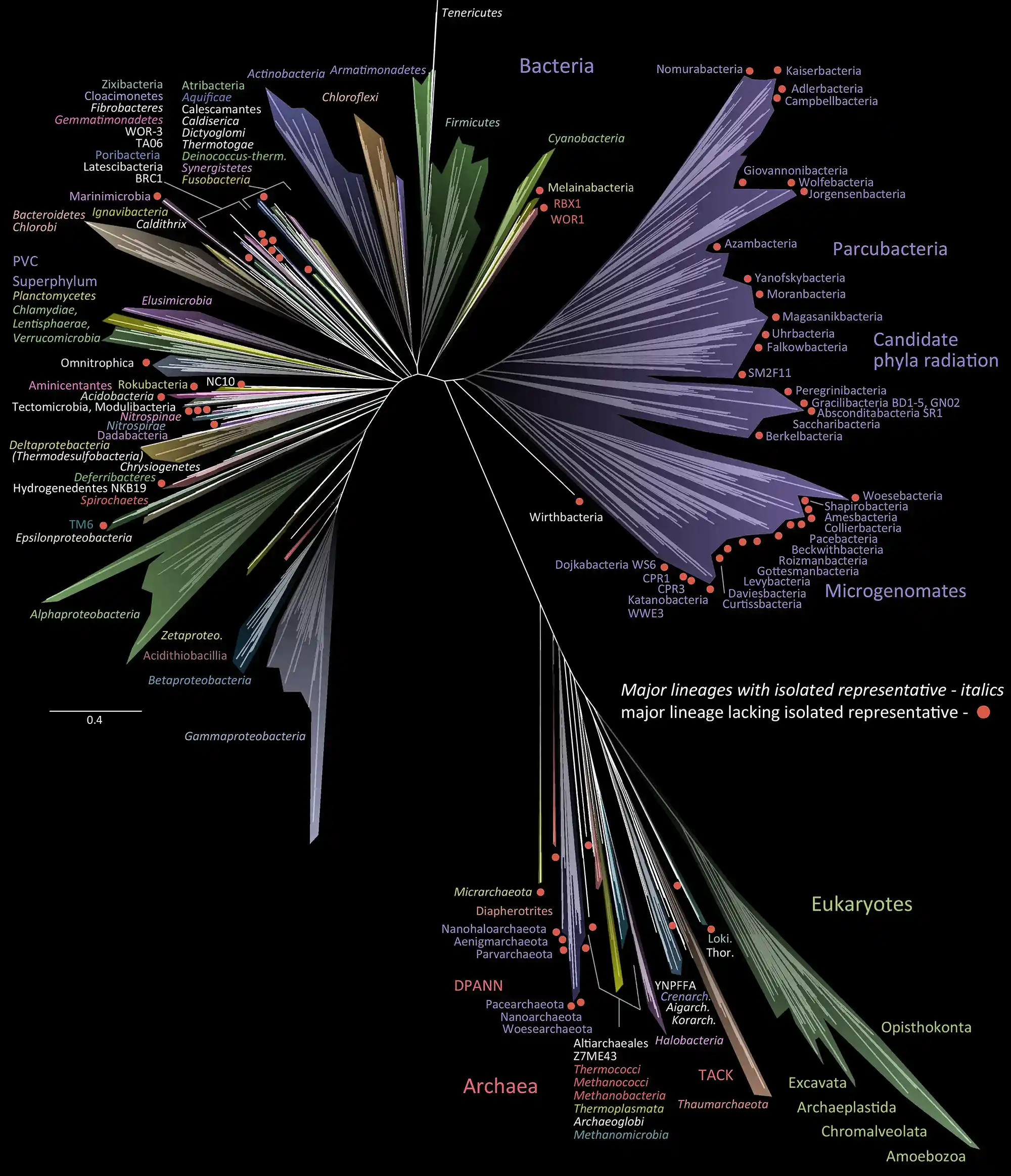
Natura Biota indikerar här naturens biom, det vill säga allt liv som existerar på jorden. Det yttersta ursprunget är inte klarlagt, men hypotetiseras bestå av den senaste universella gemensamma förfadern (LUCA), som alla i dag levande organismer stammar från. LUCA utgör däremot inte det första livet, på samma sätt som Y-kromosoms-Adam och mtDNA-Eva inte utgjorde de första människorna.
Archaea (-3500 Ma) är en domän som har utvecklats ur LUCA, och den omfattar organismer som redan är välutvecklade biologiska fabriker, sprungna ur tidig biologisk evolution och än tidigare kemisk evolution under abiogenesen, den emergenta process i vilken autokatalytiska system med självreplikering av RNA-strängar inkapslade i en protocell med ett skyddande lipidhölje uppstod. Det är ytterst en fråga om effektiv elektrontransport för att finna minimal energinivå för något molekylärt tillstånd.
Arkéer skiljer sig från bakterier (Bacteria) genom eterbindning i membranlipider samt mer komplex transkription av RNA till mRNA och slutligen proteiner. Arkéer och bakterier hör till prokaryoter, organismer med cellkärna utan cellmembran, mitokondrier eller andra organeller.
Människan är dock varken en arké eller en bakterie, utan en eukaryot (Eukaryota), en tredje domän som kännetecknas av mer komplexa celler samt en cellkärna med cellmembran. De tre domänerna kan formellt skiljas ut genom hur ribosomalt rRNA verkar i cellerna.
Eukaryota (-1850 Ma) antas ha uppstått genom symbiogenes av bakterier och arkéer, nämligen sammanslagning av Bacteria Proteobacteria Alphaproteobacteria och Archaea Proteoarchaeota Eukaryomorpha (Asgard). Fria prokaryoter har således fångats i en gemensam cell och där förslavats i vidare utveckling mot mer komplexitet, varvid α-Proteobacteria har blivit en mitokondrie. Vidare endosymbios med cyanobakterier har på samma sätt gett upphov till kloroplaster, organeller förmögna till fotosyntes i alger och växter.
Eukaryoter kännetecknas av förmåga till asexuell reproduktion i mitos och sexuell i meios, samt av flercellighet med epigenetiskt bestämda celltyper i olika slags vävnader. Djur, växter och svampar är de vanligaste rikena i domänen, med en rest i protister för organismer som är svårare att klassificera.
I det vidare fylogenetiska utvecklingsträdet finner man ett antal klader eller grenar som representerar distinkta funktioner, till exempel Orthokaryota, som bland annat definieras av ortogonala centrioler och en staplad golgiapparat, samt Neokaryota, som avskiljer basala encelliga heterotrofa eukaryoter i Excavata från den gren som leder till djur, växter och svampar.
Neokaryoter förgrenas i sin tur i å ena sidan Bikonta (organismer med celler med två flageller), som leder vidare till Archaeplastida med alger och landväxter, och å den andra Scotokaryota, som leder till djur och svampar. Vidare förgrening av Scotokaryota ges av Podiata, som skiljer ut andra organismer utanför linjen till djur och svampar.
Unikonta (organismer med celler med en flagell, -1400 Ma) följer ur Podiata, och har en gren i Amoebozoa, en återvändsgränd som definierar amöbor och protister, men också en som leder vidare till mer avancerade organismer i Obazoa. Unikonter skiljer sig i övrigt från bikonter genom förekomsten av en tredubbel genfusion.
Obazoa (-1301 Ma) förgrenar sig dels i Breviata och Apusomonadida, två klasser av zooflagellater utan vidare förgrening, och dels i den viktiga kladen Opisthokonta (-1300 Ma). Opistokonter kännetecknas av att cellerna har en enskild flagell i bakre ändan, medan andra eukaryoter kan ha flageller i främre ändan. Har celler hos människor verkligen en sådan flagell? Ja, i form av spermier, innan den fortsatta kroppsplanen manifesteras enligt senare evolutionär utveckling.
Opistokonterna har två utvecklingslinjer, där Holomycota leder till svampriket, och Holozoa (-1100 Ma) till djur och en del encelliga organismer som choanoflagellater. Även Filozoa (-1000 Ma) och Choanozoa (-950 Ma) skiljer ut djur från en del andra organismer i den vidare utvecklingen, till exempel choanoflagellater.
Animalia (-665 Ma) eller riket av djur följer ur Choanozoa. Typiskt för organismer i riket är förmåga till rörlighet, heterotroft näringsupptag, aerobisk respiration och sexuell fortplantning. Sådana organismer möjliggjordes genom tillväxt av syre i atmosfären under Kryogenium -720/-635 Ma som ett resultat av cyanobakteriers respiration.
Ett underrike till Animalia utgörs av Eumetazoa (-635 Ma) som omfattar alla flercelliga djur med äkta cellvävnad, till skillnad från svampdjur i systergrenen Porifera. I Eumetazoa är Parahoxozoa (-605 Ma) vidare en monofyletisk klad som definierar organismer med så kallade hox-gener för uttryck av kroppsplanen. Kladen står i kontrast till Ctenophora eller kammaneter, ryggradslösa djur som saknar sådana genuttryck.
I den vidare taxonomin följer tvåsidiga djur i Bilateria (-560 Ma), organismer som har två i stort sett symmetriska kroppshalvor. De flesta djur har sådan symmetri, förutom organismer i systerkladen Cnidaria och Placozoa, kammaneter och svampdjur. Symmetrin behöver inte vara fullständig, varför enstaka organ som hjärta kan förekomma ensidigt.
Den tvåsidiga strukturen innebär i regel att bilaterala organismer har öppningar i främre respektive bakre ändan för tillförsel av näring och utsöndring av exkrementer. Öppningarna bildas under gastrulation i embryonalt stadium, då en skålformad mage bildar centrum av ett matsmältningssystem. Öppningarna bildas inte samtidigt, utan sker konsekutivt. Existensen av en väldefinierad främre ända främjar samtidigt framväxten av ett huvud med sensoriska organ och ett nervsystem, och småningom även en hjärna.
Deuterostomia (-540 Ma) betecknar ett överfylum av sådana organismer i vilka den ursprungliga öppningen fungerar som anus, medan Protostomia (-540 Ma) utgör ett systerfylum med organismer i vilka den ursprungliga öppningen fungerar som mun. De båda samlas i Nephrozoa (-558 Ma), som avskiljer fylumet Xenacoelomorpha av organismer utan verkligt matsmältningssystem.
Till protostomerna hör de flesta ryggradslösa djur likväl som leddjur i form av insekter och kräftdjur. Deuterostomerna utvecklas å ena sidan i sjögurkor, sjöstjärnor och andra ryggradslösa tagghudingar i Ambulacraria, och å den andra i ryggsträngsdjur i fylumet Chordata (-535 Ma) med bland annat en ryggsträng och en svansstruktur i förlängning av anus. Ryggsträngsdjuren framträder under den kambriska explosionen, med snabb evolution i avancerat liv.
Ryggsträngsdjuren har en förgrening i kladen Olfactores (-535 Ma) respektive Cephalochordata, där den senare främst utgörs av enkla lansettfiskar. Olfactores indelas vidare i underfylumet Tunicata av manteldjur omfattande sjöpungar och andra akvatiska organismer, samt i kraniedjur i Craniata (-535 Ma).
Kraniedjuren är nära nog synonyma med ryggradsdjuren i underfylum Vertebrata (-520 Ma), som kännetecknas av organismer med ett skyddande hölje runt ryggsträngen. Höljet bildar ett endoskelett, som ger organismen struktur och bättre rörelseförmåga. Pirål är dock ett exempel på ett kraniedjur som inte är en vertebrat, varför en distinktion kan motiveras.
Gnathostomata (-462 Ma) eller käkmunnar utgör ett infrafylum av den förkrossande majoriteten av ryggradsdjur, med pansarhaj som en tidig företrädare och nutida ben- och broskfiskar som moderna. Utvecklingen av däggdjur och människor har således gått via fiskar med gälar och fenor. En underavdelning är Eugnathostomata av verkliga käkmunnar, till skillnad från primitiva pansarhajar.
Verkliga käkmunnar har en vidare indelning i Euteleostomi eller Osteichthyes (-430 Ma), det vill säga benfiskar, som alltjämt är den största klassen av ryggradsdjur. En parallell klass (relativt fiskar) utgörs av bronsfiskar i Chondrichthyes, till vilken hajar och rockor hör, samt den utdöda klassen Acanthodii av hajliknande fiskar med romboida fjäll.
I Euteleostomi återfinns de strålfeniga fiskarna i Actinopterygii, som omfattar nästan alla nu levande fiskar. Den parallella underklassen av lobfeniga fiskar i Sarcopterygii (-423 Ma) har i nutid lungfiskar som representanter. Ur de lobfeniga fiskarna stammar dels numera utdöda kvastfeniga fiskar i Rhipidistia (-416 Ma) och vidare i Tetrapodomorpha (-409 Ma), fiskar som ur fenor utvecklade fotliknande extremiteter för att mer effektivt kunna fånga byten i strand- och bottennära miljöer.
En vidare indelning av Tetrapodomorpha har vi i Eotetrapodiformes (-400 ma), Elpistostegalia (-385 Ma) och Stegocephalia (-375 Ma), i allt finare utveckling mot amfibieliknande egenskaper i landryggradsdjuren Tetrapoda (-370 Ma). Fyrfotingarna utvecklade dock sina extremiteter i vattenmiljö innan de nyttjades på land.
Tetrapoderna utvecklades i å ena sidan Batrachomorpha och Amphibia, som inte leder till människan, och å andra sidan Reptiliomorpha (-340 Ma) av reptilliknande groddjur i en process som ledde till intern fertilisering i ägg som lades på land snarare än i vatten, där den senare egenskapen är kännetecknande för Amniota (-330 Ma).
Amniota omfattar kräldjur (inklusive fåglar) i Reptilia och synapsider i Synapsida (-312 Ma), men alltså inte groddjur i Amphibia. Synapsider eller stamdäggdjur är den linje som leder till oss, och vi är därför inte släkt med kräldjuren utan utgör en parallell utveckling. Kännetecknande för synapsider är tinningsöppningar i kraniet, som ger dels bättre fäste för käkmuskler och dels mindre vikt. En differentierad tandrad med specialiserade tandfunktioner kan också skönjas.
Synapsiderna karakteriseras vidare av förmåga till upprätt kroppshållning, endoterm reglering av kroppstemperaturen (varmblodighet), en protopäls, samt mjölkkörtlar för att fukta ägg utan kalciumskal, särskilt i senare utvecklingar som Eupelycosauria, Metopophora, Haptodontiformes, Sphenacomorpha, Sphenacodontia, Pantherapsida och Sphenacodontoidea (-300 Ma), som främst definieras genom varierande proportioner i kraniets ben och tänder.
Ordningen Therapsida (-275 Ma) utgör ättlingar till synapsiderna, och kännetecknas bland annat av lemmar under kroppen snarare än utstrålande från sidan som hos reptiler. Hår, inklusive morrhår, förekommer i senare utvecklingar, likväl som mjölktänder, tre ben i mellanörat, sju halskotor och en särskild bröstkorg. Likt synapsiderna har therapsiderna en gradvis raffinering, som klassificeras i kladen Eutherapsida (-275 Ma), Neotherapsida (-270 Ma), Theriodontia (-270 Ma) och Eutheriodontia (-265 Ma), med mer däggdjursliknande tänder.
Cynodontia (-260 Ma) – «hundtänder» – hör till en underordning av therapsider som överlevde perm–triasutdöendet vid -252 Ma, och därefter var hänvisad till att leva i skuggan av de dinosaurier som dominerade. Therapsiderna hade tidigare själv dominerat landlivet, men fick hädanefter nöja sig med ett råttliknande nattaktivt liv i mörker och under jord. Levernet gav sämre syn, men främjade bättre hörsel samt sensoriska morrhår. Cynodonterna var också först med att kunna tugga och andas samtidigt, en egenskap du förmodligen aldrig har reflekterat över.
Den vidare vägen mot däggdjur går via Epicynodontia (-260 Ma), infraordningen Eucynodontia (-250 Ma), parvordningen Probainognathia (-240 Ma) samt kladen Chiniquodontoidea (-230 Ma) och Prozostrodontia (-230 Ma), med mindre gradvisa förändringar i anatomi och funktion.
Mammaliamorpha och det snävare Mammaliaformes (-225 Ma) av däggdjursliknande organismer följer ur cynodonternas utveckling. Mjölktänder och ej ersättningsbara permanenta tänder med emalj underlättar nedbrytningen av föda för större kaloribehov vid endoterm reglering. Digivning till ungar är en typisk egenskap. Däggdjuren i klassen Mammalia (-225 Ma) kännetecknas även av ett mellangärde, ett hjärta med två kammare och förmak samt termoreglering kontrollerad av hjärnan.
Processen att förlägga fertiliseringen internt istället för i ägg fullbordas i Theriiformes (-220 Ma), som avskiljer monotremer som kloakdjur. Holotheria (-220 Ma) och Trechnotheria (-216 Ma) är två raffineringar. Cladotheria, Zatheria och Tribosphenida (-165 Ma) utvecklar moderna skulderblad och förfiningar av bakbenen samt ytterligare förändringar i tandraden.
Födsel av levande ungar uppträder därefter i Theria och Eutheria («riktiga bestar») (-161 Ma), med en näbbmusliknande och trädlevande Juramaia sinensis som första kända representant.
Högre däggdjur i infraklassen Placentalia, det vill säga moderkaksdjur, uppträder i fossilen från -66 Ma, då dinosauriernas herravälde är till ända, men de uppstod troligen tidigare. De radieras emellertid snabbt då nya nicher blir tillgängliga, och däggdjuren blir snabbt dominerande. Utan det konstanta hotet från dinosaurier ökar däggdjuren även snabbt i storlek och blir dagaktiva.
Magnordningen Boreoeutheria (-65 Ma) innehåller dels kladen Laurasiatheria av placentadäggdjur på den laurasiska kontinenten, numera omfattande rovdjur, fladdermöss, näbbmöss, valar och hovdjur, och dels överordningen Euarchontoglires (-65 Ma) av gnagare, hardjur, spetsekorrar, pälsfladdrare och – primater. De tre sista djurgrupperna samlas i den snävare storordningen Euarchonta (-65 Ma) av «sanna förfäder».
Mirordningen Primatomorpha (-65 Ma) reducerar gruppen ytterligare till att omfatta dels pälsfladdrare i ordningen Dermoptera, dels primater i ordningen Primates (-55 Ma). Primaterna indelas i sin tur i underordningen Strepsirrhini eller lägre primater, samt i Haplorhini (-50 Ma) eller högre primater. De högre primaterna är dagaktiva och förlitar sig mer på syn än lukt. De lägre primaterna kännetecknas istället av våta näsor och därför bättre luktsinne.
Haplorhini har en indelning i infraordningen Tarsiiformes och dess systerordning Simiiformes (-40 Ma). Den senare förgrenas i dels brednäsor i parvordningen Platyrrhini (-31 Ma), som uteslutande har sina habitat i den Nya världen, dels smalnäsor i Catarrhini (-35 Ma) i den gamla. Vi människor är smalnäsor med parallella näsborrar och en motsättlig tumme för bättre gripförmåga, medan brednäsorna aldrig lämnade träden.
Smalnäsorna har en vidare indelning i markattartade apor i Cercopithecidae samt i överfamiljen Hominoidea (-21 Ma) av människoartade apor. Hominoidea kännetecknas av större rörelseförmåga till följd av flitigt svingande i träd. En vidare förgrening görs i gibboner i Hylobatidae samt i familjen Hominidae (-17 Ma) omfattande gorillor, schimpanser, orangutanger, bonobo och människor.
Underfamiljen Homininae (-13 Ma) skiljer ut gorillor i tribus Gorillini och övriga i Hominini (-6 Ma), medan orangutangerna finns i systergruppen Ponginae. Hominini består i sin tur av subtribus Panina omfattande schimpanser i släktet Pan samt av subtribus Hominina (-4 Ma) respektive Australopithecina (-5.6 Ma).
Människans utvecklingslinje korsas av såväl Hominina som Australopithecina, och resulterar i släktet Homo (-2.8 Ma), med H. habilis (-2.1 Ma), H. erectus (-1.9 Ma) och H. heidelbergensis (-0.6 Ma) som några representanter. Kladogrammen är raka, men den egentliga utvecklingen är mer vävformad med artvisa återkopplingar.
Arten Homo sapiens uppstår emergent i Afrika vid -350 ka, vilket ska tolkas som utveckling under en tidsperiod och omfattande flera parallella linjer. Det är anatomiskt moderna människor, men de ser inte riktigt ut som oss, och de har inte heller samma beteende. Andra arter i samma släkte är H. neanderthalensis (-250 ka), H. floresiensis (-190 ka), H. luzonensis (-67 ka) samt H. denisova (-40 ka).
Arkaisk H. sapiens vidareutvecklas i och över hela Afrika under 280 ka. Senaste anfader till alla nu levande män representeras av Y-kromosoms-Adam från -275 ka, och motsvarande gemensamma urmoder av Mitokondriska Eva från -150 ka, med en obruten linje av döttrar in i vår tid.
Homo sapiens förgrenas vidare i underarten Homo sapiens sapiens, med syster i Homo sapiens idaltu (-160 ka), en nu utdöd människotyp. Klassificeringen är osäker, och en del menar att även H. neanderthalensis är en underart H. sapiens neanderthalensis.
Vid -70 ka vandrar arkaisk H. sapiens ut ur Afrika, likt tidigare H. erectus och H. heidelbergensis, där den senare småningom övergick till H. neanderthalensis i Europa. H. sapiens, som också stammar från H. heidelbergensis, stöter på sin kusin H. neanderthalensis i Levanten, förmodligen under fientliga former.
Men det sker även genetiskt utbyte mellan de två homininerna, frivilligt eller ej, vilket resulterar i en hybridform. I dag har ickeafrikanska människor alltjämt 2–7 % DNA från neandertal, omfattande runt en femtedel av neandertals fullständiga genom. Andelen var tidigare långt högre, men har rensats bort successivt.
Vidare migration från Levanten till Europa sker från -50 ka under senaste glaciala maximum, med ytterligare beblandning med neandertal. Motsvarande process sker även i Östasien, fast med H. denisova, som är en östlig utveckling av neandertal.
Den hybrid som blir resultatet av denna utveckling definierar den beteendemässigt moderna människan, benämnd tidig europisk modern människa (EEMH), och kännetecknas av en explosion i abstrakt tänkande, symboliskt beteende, teknisk miniatyrisering, kulturell utveckling med mera, representerat av istidskonsten i allt från aurignaciens venusstatyetter och musikinstrument till solutréens och magdaleniens grottmålningar i Altamira och Lascaux. En liknande process äger rum i Asien, och småningom diffunderar arvet även till Afrika i tillämpliga delar.
Man kan av utvecklingen sedan utvandringen ur Afrika dela in det moderna människosläktet i ett antal underarter, även om det av politiska skäl inte låter sig göras i praktiken. Till det inledande stamträdet kan man då foga Homo sapiens eurasica för västeuraser, och i mitt fall skulle man även kunna lägga till WHG/EEF/ANE I-M253 för att slå fast ett stridsyxearv av istidens västliga jägare–samlare (WHG), tidiga europeiska bönder (EEF) från Anatolien vid civilisationens gryning, samt äldre nordeuraser (ANE) i form av expanderande indoeuropéer vid -5 ka. I-M253 är en haplogrupp (Y-kromosom) som är ursprunglig för Europa i en mutation som ägde rum -5 ka och som är typisk för Sverige och Skandinavien.
Givet ett stridsyxearv kan man även (i princip) härleda en sedvanlig stamtavla från -2 900 och framåt, det vill säga 4 900 år eller 196 generationer av män och kvinnor som har fört sitt arv vidare. I genetisk mening finns ingen evolutionär utveckling att tala om under den tidsrymden, och praktiskt taget inget unikt för dessa individer finns kvar i den överförda arvsmassan – den späds ut med 50 % per generation under rekombination av DNA i könsceller. Det enda som sprids är i regel det gemensamma och det bestående för denna grupp av individer.
Evolutionen vilar dock aldrig, och småningom kommer en ny gren att krystalliseras ut ur Homo sapiens och färdas vidare mot ytterligare raffinering och anpassning, på samma sätt som har skett oräkneliga gånger under fyra miljarder år av oförtröttlig biologisk evolution. Om ytterligare 50 k år har vi möjligen en klart synlig förändring.